The history of Central Asia and Anatolia make it clear that Turkic and Kurd speaking people were late comers in the History of these areas. Prior to their migration into many parts of these areas Dravidian, Kushite (the Hattic, Mittanni, Kaska and etc. speaking groups) and Sumerian speaking people left numerous inscriptions in non-Turkic and European languages, as well as Egyptian and Hittite documents testifying to the absence of Turkic speaking nationalities inhabiting the area. All of these languages are genetically related to Tamil, like Sumerian and Elamite.
The history of the Turkish languages and the earlier presence of Sumerian and Tamil speaking people in Central Asia and Anatolia, explains any relationship you might find between Turkish and the Sumerian and Tamil languages. Moreover, the fact that the Greeks were in India when Panini wrote his grammar of Sanskrit, explains the Greek relationship with Sanskrit.; and the Elamite speaking Achaemenids (and Persians)also ruled India for years and thus placed Persian and Elamite Arya and Hindi Arya in intimate contact for hundreds of years.
The relationship between the Hindi and Iranian languages is best explained by history rather than some ancient Indo-Iranian group of languages.
Achaemenid records make it clear that by 559 BC, Persian speakers were ruling the Hindus. Given the fact that their is historical evidence of Persians ruling Hindus can explain the relationship between the speakers of these languages instead of the Indo-European hypothesis. These Persians also ruled the Greeks and the Dravidian speaking Lycians in Europe.
The Achaeminids spoke Persian and Elamite. Since they were rulers of the Indians would have been natural for the Hindus to adopt many Persian terms and vice versa during a period of bilingualism in North India. Since the Elamite and Dravidian speakers spoke genetic languages there was little need for a lingua franca to allow communication between the diverse groups under Achaeminid rule.
King Darius

King Darius (521-486) made it clear he was an Arya "nobleman". The first use of Arya had nothing to do with race. Darius , in an inscription at Naghsh-Rostam (near Shiraz in present-day Iran) noted that:
- "I am Darius the great king, a Persian, son of a Persian, an Aryan, and having Aryan lineage."
Darius later conquered Macedonia. This led to Achaeminid people speaking Elamite and Persian may have contributed lexical items to ancient Ionian (Greek). Since we have historical evidence of a close relationship between Hindus and Persians by the 6th Century BC, there is no need to claim that the relationship between Indo-Iranian began in 1200 BC,when the historical evidence indicates interaction between these groups by 589 BC, not 1200 BC. As a result of the Persians living in Iran, up until the Achaeminid Empire, there is no way anyone can claim that the Indo-Iranian homeland was in India. There was no Indo-Iranian homeland, the relationship between these languages was probably the result of the Achaeminid rule of India.
The Persians also ruled the Greeks. The Greeks later conquered India, and Panini mentions Greeks in his grammar of Sanskrit. This suggest that Greeks lived in large numbers in India at this time.The fact that the Greeks, Hindus and Persians lived in intimate contact for hundreds of years naturally led to the adoption of many terms by the Greeks and Hindus of Persian, and later the adoption of many Greek terms by the Hindus. These states of bilingualism in North India, explains why the Indo-Iranian languages form one family , and are linked to the Indo-European languages via Greek.
The Harappans spoke a Dravidian language, Indo-Iranian probably originated after 589 BC. This is made clear by Darius in the Behistun inscription where he claims that he was the first to write in the Ariya language. Darius'- evidence for the first writing of Ariya, indicates that the idea of the continuity of Hindu civilization in India is a myth. The original inhabitants of India spoke Dravidian languages. Over time, the Dravidians were forced to adopt Hindi and other Indo-Iranian languages, yet remnants of these Dravidians in North India remain.
This is why we find no evidence of the Vedic language until the Naga (Ethiopians) invented Sanskrit. It also explains the variations in the Vedic and Avestan manuscripts, which in the case of the later group date back only to 1288 AD.
The tradition of writing in North India date to the Achaeminids, and may explain the origin of Brahmi. The fact that Brahmi has signs that relate to the Harappan writing may be the result of the fact that the Elamites of the Achaeminid Empire were familiar with the writing system of the Dravidians, and the Naga (Ethiopians) who used a system of writing similar to Phonesian.
The Dravidians have their own tradition of writing.It would appear that the Dravidians introduced writing to the Indus Valley. They continued to use this writing on their pottery in South India and later punchmarked coins. This is supported by the discovery of writing in South India dating back to before 600 BC.
The history of contact between Iranian and Indian speakers during Achaemenid rule , would explain the Indo-Iranian relationship, not the existence of a Proto-Indo-Iranian homeland in India. This history of Turkic, Persian, Sumerian, Elamite, Tamil, Ethiopic (/Naga)and Hindi speaking people living in diverse North Indian communities, is the most logical explanation of the relationships that exist between and among these languages. The history of linguistic contact between the speakers of these languages make it clear that the Harappans were not Indo-Aryan
speakers. This would place the origin of the major Vedic and Avestan text back to maybe 800 BC, and more than likely 600-500 BC not the 1200 BC or earlier date assigned these text by some researchers. Let's not forget that some researchers claim that most editions of the Avestan, date back to an original copy of this text dating only to 1200 AD.
In summary India was not the home land of the Indo-Iranian family of languages.The linguistic relationship between Persian and Greek result from the rule of these areas by the Achaeminid and later Greek rulers of India. This may explain why the Achaeminids depicted the Nubians (of Africa), the Hindus and King Darius with Africoid features.
The ability to explain the relationship of Sanskrit to Greek, and the Indo-Iranian linguistic relationship due toPersian/ Elamite and Hindi contact, resulting from the historical connections between the speakers of these languages and bilingualism within North-India and Afganistan. This hypothesis supports the view that the Indo-European connection to Indian languages goes back to the Greek rule of India, not some hypothetical date millennia ago.
Africans carry haplogroup R1a.
In India the Dravidian people carry the R1a haplogroup The Dravidian people of India originally lived in Middle Africa and belonged to the Proto-Saharan Civilization.
The Proto-Saharan civilization was situated in the Proto-Sahara, which includes Cameroon.
.
 .
In Cameroon we find carriers of R1a.
.
In Cameroon we find carriers of R1a.
In addition to carriers of R1a in Cameroon; the Dravidian languages are still spoken today in Cameroon see: https://www.youtube.com/watch?v=vWyAYGlFZjkhttps://www.youtube.com/watch?v=vWyAYGlFZ jk
See:
- Gemma Berniell-Lee,
- Francesc Calafell,
- Elena Bosch,
- Evelyne Heyer,
- Lucas Sica,
- Patrick Mouguiama-Daouda,
- Lolke van der Veen,
- Jean-Marie Hombert,
- Lluis Quintana-Murci,
- and David Comas
Genetic and Demographic Implications of the Bantu Expansion: Insights from Human Paternal LineagesMol Biol Evol (2009) 26 (7): 1581-1589 first published online April 15, 2009doi:10.1093/molbev/msp069
Central Asia was called Kush by the ancient people. The Armenians made it clear that the ancients called Persia, Media,Elam Aria, and the entire area between the Tigris and Indus rivers Kush.Bardesones, writing in his Book of the Laws of Countries, in the 2nd Century said that the "Bactrians who we called Qushani (or Kushans)".The Armenians, called the earlier Parthian: Kushan and acknowledged their connection with them. Homer, Herodotus, and the Roman scholar Strabo called southern Persia AETHIOPIA. The Greeks and Romans called the country east of Kerma: Kusan.
First, I would like to make it clear that the probable language of the Kushana was Tamil. According to Dravidian literature, the Kushana were called Kosars=Yakshas=Yueh chih/ Kushana. This literature maintains that when they entered India they either already spoke Tamil, or adopted the language upon settlement in India. In pinyin Yueh chih is pronounced: Yuezhi
The Kushana and the Yueh chih/ Yuezhi were one and the same. In addition to North Indian documents the Kushana-Yueh chih association are also discussed in Dravidian literature. V Kanakasabhai, The Tamils Eighteen hundred years ago, note that in the Sanskrit literature the Yueh chih/ Yuezhiwere called Yakshas, Pali chroniclers called them Yakkos and Kosars< Kushana.
Many of the Yueh people were Dravidian speakers. The Yueh people were also called Yuehchih or Kuishuang (Kushana). In ancient times the Yueh chihs controlled Central Asia and much of China until the first century BC. In the Pali Chronicles, the Ramayana and Matsya, the Yuehchih were called Yakshas or Kosar. The Yueh of North China established Xia. According to the Yi Xia Dong Xi Shuo, by Fu Ssumein, the li Qiang (Black Qiang) of Shang were united with the Yueh people of southwest China.
.
 .
The Yuezhi allegedly arrived in India during the 2nd century BC. He makes it clear that the Yuezhi / Kushana as noted on their coins worshipped Siva as seen on the coins of Kanishka. This is why we have a coin of a Kushana king from Taxila, dated to AD 76 that declares that the king was maharaja rajatiraja devaputra Kushana "Great King, King of kings, Son of God, the Kushana".
The term Tochara has nothing to do with the Yuehchih, this was a term used to describe the people who took over the Greek Bactrian state, before the Kushana reached the Oxus Valley around 150 BC . There is no reason the Kushana may not have been intimately familiar with the Kharosthi writing at this time because from 202BC onward Prakrit and Chinese documents were written in Kharosthi.
The Kushana and the Yuezhi were one and the same. In addition to
North Indian documents the Kushana- Yuezhi association are also discussed in Dravidian literature.V Kanakasabhai, The Tamils Eighteen hundred years ago note that in the Sanskrit literature the Yueh chih were called Yakshas, Pali chroniclers called them Yakkos and Kosars< Kushana.
Some researchers believe that the Ars'i spoke Tocharian A, while Tocharian B was the "Kucha language" may have been spoken by the Kushana people. I don't know where you read that the speakers of Tocharian A were called Ars'i. These names: Ars’i and kucha, have nothing to do with ethnic groups, they refer to the cities where Tocharian text were found: Tocharian A documents were found around Qarashar and Turfan, thusly these text are also referred to as Turfanian or East Tocharian; Tocharian B documents were found near the town of Kucha, thusly they are sometimes called Kuchean or West Tocharian.
Linguist use the term Tochari to refer to these people, because they were given this title in Turkic manuscripts . As a result, the word Tochari has nothing to do with the Kushana people. The observable evidence make it clear that the terms used to label the Tocharian dialects are not ethnonyms, they are terms used to denote where the Tocharian records were found. The use of the term Ars'i does not relate to the Kushana people. The terms: Asii, Pasiani, Tochari and Sacarauli, refer to the white nomads that took Bactria away from the Greeks—not the Yuezhi .
These white nomads came from the Iaxartes River that adjoins that of Sacae and the Sogdiani .The Kushana people took over Bactria much later. It is a mistake to believe that Ars'i and Kucha were ethnonyms is understandable given your lack of knowledge about Tocharian. And I will agree that there were a number of different languages spoken by people who wrote material in Tocharian. It is for this reason that I have maintained throughout my published works on Tocharian, that this was a trade language. See: Tocharian is a Dravidian trade languagehttps://www.academia.edu/8491572/Is_Tocharian_a_Dravidian_Trade_Language
This Tocharian/Kushan language was used by the Central Asians as a lingua franca and trade language due to the numerous ethnic groups which formerly lived in central Asia". Kharosthi was long used to write in Central Asia. It was even used by the Greeks. The use of the Kharosthi writing system in Central Asia and India, would place this writing contemporaneous with the tradition, recorded by the Classical writers of Indians settling among the Kushana.
There were many people who probably used Tocharian for purposes of communication including the Kushana and the "Ars'i/Asii". They probably used Tocharian as a lingua franca. You make it clear in your last post that numerous languages were spoken in Central Asia when the Tocharian was written in Kharosthi. Most researchers believe that a majority of the people who lived in this area were bilingual and spoke Bactrian ,Indian languages among other languages. I agree with this theory, and believe that the Kushana Kings may have spoken a Dravidian language. Due to the possibility that the Kushana spoke a Dravidian language which is the substratum language of Tocharian; and
the presence of a number of different terms in Tocharian from many languages spoken in the area-led me to the conclusion that Tocharian was a trade language. The Kushana always referred to themselves as the Kushana/Gushana. The name Kushana for this group is recorded in the Manikiala Stone inscription (56BC?), the Panjtar Stone inscription of 122 AD and the Taxila Silver Scroll.
The Greeks called them Kushana in the Karosthi inscriptions, and Kocano. In the Chinese sources they were called Koei-shuang or Kwei-shwang= Kushana, and Yueh chih .
As you can see the term Kushana had been used to refer to these people long before Kujula Kadphises used the term as a personal name. This was over a hundred years after the Kushana had become rulers of Bactria. It would appear from the evidence that the nation of the Kushana was called Kusha.
In 176 B.C., the Huns fell upon the in western Gansu,defeated their army and murdered their King. This battle led to the Kushana migration into Nanshan region, and thence to Bactria and North India. (Bagchi 1955, p.4)
The Kushana first occupied Transoxiana about 160 B.C. and established themselves in the Oxus Valley (Chi 1955, p.8) They later drove the Haumavorka Indo-European Saka people, from Bactria and founded the Kushana dynasty which lasted until the 3rd century A.D.
It was Kujuula Kadphises who united the Kushana people and made them into a single nation. Kadphises conquered India as far as the Indus. His capital was Purushapura near Peshwar, in Pakistan. Later Wiima Kadphises extended Kushana rule into the Punjab.
The Kushana conquered the Sakas and Parthians and took control of an empire stretching from the Oxus river in Afghanistan, to the Ganges plains of India.. This unite under one authority the former dominions of the Indo-Greeks and the Sunga dynasts.
The greatest king of the Kushana was Kaniska. Kaniska came to power between A.D. 78-144. (Thapar 1972, p.92)
.
The Yuezhi allegedly arrived in India during the 2nd century BC. He makes it clear that the Yuezhi / Kushana as noted on their coins worshipped Siva as seen on the coins of Kanishka. This is why we have a coin of a Kushana king from Taxila, dated to AD 76 that declares that the king was maharaja rajatiraja devaputra Kushana "Great King, King of kings, Son of God, the Kushana".
The term Tochara has nothing to do with the Yuehchih, this was a term used to describe the people who took over the Greek Bactrian state, before the Kushana reached the Oxus Valley around 150 BC . There is no reason the Kushana may not have been intimately familiar with the Kharosthi writing at this time because from 202BC onward Prakrit and Chinese documents were written in Kharosthi.
The Kushana and the Yuezhi were one and the same. In addition to
North Indian documents the Kushana- Yuezhi association are also discussed in Dravidian literature.V Kanakasabhai, The Tamils Eighteen hundred years ago note that in the Sanskrit literature the Yueh chih were called Yakshas, Pali chroniclers called them Yakkos and Kosars< Kushana.
Some researchers believe that the Ars'i spoke Tocharian A, while Tocharian B was the "Kucha language" may have been spoken by the Kushana people. I don't know where you read that the speakers of Tocharian A were called Ars'i. These names: Ars’i and kucha, have nothing to do with ethnic groups, they refer to the cities where Tocharian text were found: Tocharian A documents were found around Qarashar and Turfan, thusly these text are also referred to as Turfanian or East Tocharian; Tocharian B documents were found near the town of Kucha, thusly they are sometimes called Kuchean or West Tocharian.
Linguist use the term Tochari to refer to these people, because they were given this title in Turkic manuscripts . As a result, the word Tochari has nothing to do with the Kushana people. The observable evidence make it clear that the terms used to label the Tocharian dialects are not ethnonyms, they are terms used to denote where the Tocharian records were found. The use of the term Ars'i does not relate to the Kushana people. The terms: Asii, Pasiani, Tochari and Sacarauli, refer to the white nomads that took Bactria away from the Greeks—not the Yuezhi .
These white nomads came from the Iaxartes River that adjoins that of Sacae and the Sogdiani .The Kushana people took over Bactria much later. It is a mistake to believe that Ars'i and Kucha were ethnonyms is understandable given your lack of knowledge about Tocharian. And I will agree that there were a number of different languages spoken by people who wrote material in Tocharian. It is for this reason that I have maintained throughout my published works on Tocharian, that this was a trade language. See: Tocharian is a Dravidian trade languagehttps://www.academia.edu/8491572/Is_Tocharian_a_Dravidian_Trade_Language
This Tocharian/Kushan language was used by the Central Asians as a lingua franca and trade language due to the numerous ethnic groups which formerly lived in central Asia". Kharosthi was long used to write in Central Asia. It was even used by the Greeks. The use of the Kharosthi writing system in Central Asia and India, would place this writing contemporaneous with the tradition, recorded by the Classical writers of Indians settling among the Kushana.
There were many people who probably used Tocharian for purposes of communication including the Kushana and the "Ars'i/Asii". They probably used Tocharian as a lingua franca. You make it clear in your last post that numerous languages were spoken in Central Asia when the Tocharian was written in Kharosthi. Most researchers believe that a majority of the people who lived in this area were bilingual and spoke Bactrian ,Indian languages among other languages. I agree with this theory, and believe that the Kushana Kings may have spoken a Dravidian language. Due to the possibility that the Kushana spoke a Dravidian language which is the substratum language of Tocharian; and
the presence of a number of different terms in Tocharian from many languages spoken in the area-led me to the conclusion that Tocharian was a trade language. The Kushana always referred to themselves as the Kushana/Gushana. The name Kushana for this group is recorded in the Manikiala Stone inscription (56BC?), the Panjtar Stone inscription of 122 AD and the Taxila Silver Scroll.
The Greeks called them Kushana in the Karosthi inscriptions, and Kocano. In the Chinese sources they were called Koei-shuang or Kwei-shwang= Kushana, and Yueh chih .
As you can see the term Kushana had been used to refer to these people long before Kujula Kadphises used the term as a personal name. This was over a hundred years after the Kushana had become rulers of Bactria. It would appear from the evidence that the nation of the Kushana was called Kusha.
In 176 B.C., the Huns fell upon the in western Gansu,defeated their army and murdered their King. This battle led to the Kushana migration into Nanshan region, and thence to Bactria and North India. (Bagchi 1955, p.4)
The Kushana first occupied Transoxiana about 160 B.C. and established themselves in the Oxus Valley (Chi 1955, p.8) They later drove the Haumavorka Indo-European Saka people, from Bactria and founded the Kushana dynasty which lasted until the 3rd century A.D.
It was Kujuula Kadphises who united the Kushana people and made them into a single nation. Kadphises conquered India as far as the Indus. His capital was Purushapura near Peshwar, in Pakistan. Later Wiima Kadphises extended Kushana rule into the Punjab.
The Kushana conquered the Sakas and Parthians and took control of an empire stretching from the Oxus river in Afghanistan, to the Ganges plains of India.. This unite under one authority the former dominions of the Indo-Greeks and the Sunga dynasts.
The greatest king of the Kushana was Kaniska. Kaniska came to power between A.D. 78-144. (Thapar 1972, p.92)
 Kaniska ruled an empire extending from Central Asia, to Varansi in the Ganges Valley. He supported the arts and repaired many Kushana monuments and cities.
Kaniska had two capitals. The capital in Central Asia was Bergraam or Kupura in Afghanistan, while in India the capital was established at Muthura.
The Kushana were not Vedic worshippers. As among the Egyptians and Nubians, the Kushana raised past kings to the status of "gods", and they dedicated temples to them.
The Kushana were great patrons of the Buddhists. They supported the Mahavana (Great Vehicle) school of Buddhism. Under the Kushana the Buddha, was depicted in the form of the Muthuras school. These Muthura school Buddhas had strong negroid features.
The Kushana king was called the raja or Maharajatiraja "king of Kings".
Another famous Kushana king, Kujula imitating the Roman denares (coins) was the first Asians to circulate coins in central Asia. It was Kaniska, who first put Buddha on Indian coins.
The Kushana made fine sculptures and engraved beautiful carved sheets of ivory. Their plaques are some of the finest art pieces in India.
.
Kaniska ruled an empire extending from Central Asia, to Varansi in the Ganges Valley. He supported the arts and repaired many Kushana monuments and cities.
Kaniska had two capitals. The capital in Central Asia was Bergraam or Kupura in Afghanistan, while in India the capital was established at Muthura.
The Kushana were not Vedic worshippers. As among the Egyptians and Nubians, the Kushana raised past kings to the status of "gods", and they dedicated temples to them.
The Kushana were great patrons of the Buddhists. They supported the Mahavana (Great Vehicle) school of Buddhism. Under the Kushana the Buddha, was depicted in the form of the Muthuras school. These Muthura school Buddhas had strong negroid features.
The Kushana king was called the raja or Maharajatiraja "king of Kings".
Another famous Kushana king, Kujula imitating the Roman denares (coins) was the first Asians to circulate coins in central Asia. It was Kaniska, who first put Buddha on Indian coins.
The Kushana made fine sculptures and engraved beautiful carved sheets of ivory. Their plaques are some of the finest art pieces in India.
.
 .
.
 .
The Kushana were at this time in control of the Silk Road, which took Chinese goods to the West. It was also under the Kushana that Buddhism entered China. The Kushana ruled India for almost 200 years.
.
The Kushana were at this time in control of the Silk Road, which took Chinese goods to the West. It was also under the Kushana that Buddhism entered China. The Kushana ruled India for almost 200 years.

The Khoisan were probably the original North Africans. The Khoisan introduced mtDNA haplogroups and y-Chromosomes hg A1 and R1-M343 to the Berbers. .

The Khoisan are the ancestors of the Black Berbers whoes descendants probably live in Morocco and the Atlas Mountains.
The Black Berbers of the Atlas Mountains and other parts of Northwest Africa are of Sub-Saharan origin and took African mtDNA into Western Europe over 40kya. The Gibraltar Straits appears to be the most reliable route for the spread of many mtDNA haplogroups from Africa, into Europe over the past 30ky (Winters,2012), including L3(M,N) .
The Khoisan carry L1c,L1i, L2b, L3d ( Rito, et al ,2013) . The motif L3b, is widespread in western Africans. It is mainly found among populations that speak languages of the Niger-Congo family like the Mandekan.
Like most African haplogroups the control region of hg L1i include 16189,16223 and 16311, just like L3a and L3b. The mutation that connects the Khoisan to the spread of L3(M,N) is AF24. The AF24 mutation is found in LOd and among the Khoisan and Senegalese .The existence of AF24 in Senegal and Southern Africa suggest that L1c, L2b, L3d and L3e is not the result of intermarriage with Bantu immigrants , as suggested by Rito et al(2013) .
LOd is the oldest mtDNA haplogroup . This haplogroup is primarily carried by the Khoisan people (Winters,2014) . It is also found among Niger-Congo speakers in West Africa where we also find LOa in West Africa in addition to L3b.
The Cro Magnon DNA found in the ancient skeletons dates back to the Aurignacian period (Winters,2011). The Cro magnon skeletons belong to the N haplogroup.
The Cro Magnon skeletons carried N1a,N1b,N1c and N* (Winters, 2010,2011). It is characterized by motifs 00073G,10873C, 10238T and A4CC between nucleotide positions 10397 and 10400. Most of the skeletons carried hg N*.
It is obvious that L3 (M,N) had expanded into Europe prior to the Neolithic.
.

.
Frigi et al (2010), in Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations noted that: “Our results also point to a less ancient western African gene flow to Tunisia involving haplogroups L2a and L3b. Thus the sub-Saharan contribution to northern Africa starting from the east would have taken place before the Neolithic. The western African contribution to North Africa should have occurred before the Sahara’s formation (15,000 BP)”.
This would explain why Pericot and Dominguez (2005) found evidence of hg L3 at ancient Iberian sites. Luis Pericot was sure that the populations associated with the Gravettian (32kya) and Soultrean (23kya) cultures were phylogenetically Sub-Saharan African (Dominguez,2005). Dominguez (2005) found that the lineages recovered from ancient skeletons associated with these cultures belonged to the African lineages L1b,L2 and L3. Almost 50% of the lineages from the Abauntz Chalcolithic deposits and Tres Montes, in Navarre are the Sub-Saharan lineages L1b,L2 and L3.
Berber and Khoisan Y-Chromosome
The archaeological data support a migration of probable Khoisan migration from Southern Africa to North Africa. You have to remember that many populations have settled Morocco, so their might not always be a one-to-one correspondence between contemporary Khoisan haplogroups and haplogroups found among contemporary Berbers in Morocco and the Atlas mountains. But given the geography, we would expect to see elements of Khoisan relic population genes among Atlas Mountain Berbers.
quote:
Originally posted by Troll Patrol # Ish Gebor:

Figure S1. Map Showing Location of the Population Samples Considered in This Study
Populations are represented by circles and numbered as in Table S5. Sectors within circles are proportional to the frequency of haplogroup A1a (green), A1b (red) and A2-T (black). Green asterisks indicate countries were haplogroup A1a has previously been observed.
See:
Table S1. Haplogroup Affiliation of the Seven Chromosomes that Were Re-sequenced
Table S5: Populations considered for the mutations defining major clades A1b, A1a and A2-T.
http://www.cell.com/cms/attachment/1088206/8032906/mmc1.pdf
The Berbers and Khoisan also carry similar R1 haplogroups. The Khoisan in South Arica carry R1-M343 (Schlebusch,,2010). Berbers at Zenata and Oran also carry R1-M343 (Bekada, et al, 2010).
The hominids and tool kits common to South Africa also appear in Morocco. It is no secret that the earliest Y-chromosome haplogroups have been found in Morocco and among the Khoisan. These haplogroups belong to hg A (M91); among the Moroccans we find A1b and A1a.The Khoisan mtDNA was named originally L1a,L1d and L1k, these clades are called LoD and LoK today.
Morocco has yielded impotant new data on African prehistory. Here we find a complex and rich set of early hominids from Jebel Irhoud, Dar es-Soltan and Contrebandiers Cave.
A pan-African Middle Stone Age (MSA) culture existed that united South Africa and Morocco.The Moroccan tools are Levallois technology and Mousterian industries were used in South Africa,North Africa and western Eurasia. Dibble et al (2013) has shown that Pan-African industries included cognate scrapers, Levallois tools, Nassarius beads and engraved ostrich shells. The Moroccans and South Africans shared Levallois tools and the use of ochre, bone tools and ostrich shells. Bouaouggar et al (2007) has shown how the shell beads from Grotte des Pigeons (Taforalt,Morocco) and South Africa's Blombos Caves. The archaeological evidence is clear the Khoisan in Morocco and South Africa shared behavioral , cognitive and technological styles. The major behavioral indicators shared by the Moroccan and South African Khoisan was mining,beads, blades, ochre and bone tools between 200-40kya. It is important to note that Moroccan tools are Levallois technologies and Mousterian industries used in North Africa and Western Eurasia. We also should note that Neanderthals also used Mousterian tools.
The Boule and Vallois research makes it clear that the Bushman expanded across Africa on into Europe via Spain as the Grimaldi people. This makes it clear that the Bushman/Khoisan people were not isolated in South Africa. The Khoisan people carry the haplogroup N. The Hadza are Bushman they carry haplogroup N.

The Aurignacian civilization was founded by the Cro-Magnon people who originated in Africa. They took this culture to Western Europe across the Straits of Gibraltar. The Cro-Magnon people were probably Bushman/Khoi.
There have been numerous "Negroid skeletons" found in Europe. Marcellin Boule and Henri Vallois, in Fossil Man, provide an entire chapter on the Africans/Negroes of Europe Anta Diop also discussed the Negroes of Europe inCivilization or Barbarism, pp.25-68. Also W.E. B. DuBois, discussed these Negroes in the The World and Africa, pp.86-89. DuBois noted that "There was once a an "uninterrupted belt' of Negro culture from Central Europe to South Africa" (p.88).
Boule and Vallois, note that "To sum up, in the most ancient skeletons from the Grotte des Enfants we have a human type which is readily comparable to modern types and especially to the Negritic or Negroid type" (p.289). They continue, "Two Neolithic individuals from Chamblandes in Switzerland are Negroid not only as regards their skulls but also in the proportions of their limbs. Several Ligurian and Lombard tombs of the Metal Ages have also yielded evidences of a Negroid element.
Since the publication of Verneau's memoir, discoveries of other Negroid skeletons in Neolithic levels in Illyria and the Balkans have been announced. The prehistoric statues, dating from the Copper Age, from Sultan Selo in Bulgaria are also thought to protray Negroids.
In 1928 Rene Bailly found in one of the caverns of Moniat, near Dinant in Belgium, a human skeleton of whose age it is difficult to be certain, but seems definitely prehistoric. It is remarkable for its Negroid characters, which give it a reseblance to the skeletons from both Grimaldi and Asselar (p.291).
Boule and Vallois, note that "We know now that the ethnography of South African tribes presents many striking similarities with the ethnography of our populations of the Reindeer Age. Not to speak of their stone implements which, as we shall see later , exhibit great similarities, Peringuey has told us that in certain burials on the South African coast 'associated with the Aurignacian or Solutrean type industry...."(p.318-319). They add, that in relation to Bushman art " This almost uninterrupted series leads us to regard the African continent as a centre of important migrations which at certain times may have played a great part in the stocking of Southern Europe. Finally, we must not forget that the Grimaldi Negroid skeletons sho many points of resemblance with the Bushman skeletons". They bear no less a resemblance to that of the fossil Man discovered at Asslar in mid-Sahara, whose characters led us to class him with the Hottentot-Bushman group.
The Boule and Vallois research makes it clear that the Bushman expanded across Africa on into Europe via Spain as the Grimaldi people. This makes it clear that the Bushman/Khoisan people were not isolated in South Africa. The Khoisan people carry the haplogroup N. The Hadza are Bushman they carry haplogroup N.
Cro-Magnon people carried haplogroup N:
quote:
Specific mtDNA sites outside HVRI were also analyzed (by amplification, cloning, and sequencing of the surrounding region) to classify more precisely the ancient sequences within the phylogenetic network of present-time mtDNAs (35, 36). Paglicci-25 has the following motifs: +7,025 AluI, 00073A, 11719G, and 12308A. Therefore, this sequence belongs to either haplogroups HV or pre-HV, two haplogroups rare in general but with a comparatively high frequencies among today's Near-Easterners (35). Paglicci-12 shows the motifs 00073G, 10873C, 10238T, and AACC between nucleotide positions 10397 and 10400, which allows the classification of this sequence into the macrohaplogroupN,containing haplogroups W, X, I, N1a, N1b, N1c, and N*. Following the definition given in ref. 36, the presence of a single mutation in 16,223 within HRVI suggests a classification of Paglicci-12 into the haplogroup N*, which is observed today in several samples from the Near East and, at lower frequencies, in the Caucasus (35). It is difficult to say whether the apparent evolutionary relationship between Paglicci-25 and Paglicci-12 and those populations is more than a coincidence. Indeed, the haplogroups to which the Cro-Magnon type sequences appear to belong are rare among modern samples, and therefore their frequencies are poorly estimated. However, genetic affinities between the first anatomically modern Europeans and current populations of the Near East make sense in the light of the likely routes of Upper Paleolithic human expansions in Europe, as documented in the archaeological record (37).
http://www.pnas.org/cgi/content/full/100/11/6593
This suggest that haplogroup N was taken to Western Eurasia by the San people=Cro-Magnon.

In summary, the Black Berbers took African mtDNA into Western Europe over 40kya . in addition, Berbers and Khoisan continue to carry y-Chromosome haplogroups A1 and R1-M343. The Tuareg probably helped spread hg H in Europe after they invaded Europe along with other sahelians/Moors during the Islamic period.
- References:
Bekada A, Arauna LR, Deba T, Calafell F, Benhamamouch S, Comas D (2015) Genetic Heterogeneity in Algerian Human Populations. PLoS ONE 10(9): e0138453. doi:10.1371/journal.pone.0138453 http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0138453
Cruciani, F., Trombetta, B., Massaia, A., Destro-Bisol, G., Sellitto, D., & Scozzari, R. (2011). A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa. American Journal of Human Genetics, 88(6), 814–818. http://doi.org/10.1016/j.ajhg.2011.05.002
Domínguez E.F. (2005). Polimorfismos de DNA mitocondrial en poblaciones antiguas de la cuenca
mediterránea. Universitat de Barcelona. Departament de Biologia Animal, 2005 (PhD thesis).
Frigi et al. (2010). Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations, Human Biology (August 2010 (82:4).
Rito T, Richards MB, Fernandes V, Alshamali F, Cerny V, et al. (2013) The First Modern Human Dispersals across Africa. PLoS ONE 8(11): e80031. doi:10.1371/journal.pone.0080031
Schlebusch, C.M. (2010). Genetic variation in Khoisan-speaking populations from southern Africa. University of the Witwatersrand, Johannesburg (2010 PhD thesis).https://www.academia.edu/3426993/Genetic_variation_in_Khoisan-speaking_populations_from_southern_Africa
Winters,C. (2010). Origin and spread of the Haplogroup N. Bioresearch Bull (2010) 3:116-122.
Winters C.(2011). The Gibraltar Out of Africa Exit for Anatomically Modern Humans. WebmedCentral BIOLOGY 2011;2(10):WMC002319 doi: 10.9754/journal.wmc.2011.002319
Winters, C. (2014). The Hadza Are Related to the South African Khoisan. http://www.exposingblacktruth.org/the-hadza-are-related-to-the-south-african-khoisan/
Winters,C.(2012). There has been a Continuous Indigenous Sub-Saharan Presence in North Africa for 30ky. Comment: . http://olmec98.net/ContinuousEurope.pdf
.
 .
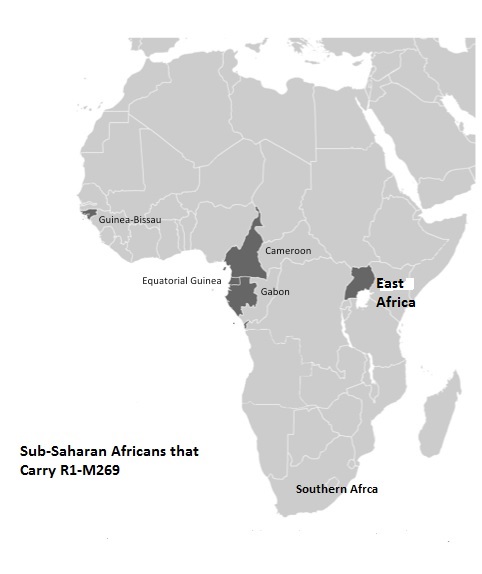
The phylogeography of R1 in Africa makes it clear that this y-chromosome is spread globally across Africa and includes the genetic structure of diverse African populations including Berber, Chadic, Cushitic, Khoisan,Pygmy, Niger-Congo, Nilo-Saharan and Semitic speaking African populations (Berniell-Lee et al, 2009; Cruciani et al, 2010; Wood et al, 2009). The fact that Dravidians carry the R haplogroup illustrate the recent introduction of R y-chromosome to Eurasia.
Abu-Amero et al (20009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of Ry-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa, especially Cameroon (Coia et al, 2005). Coia et al (2005) has revealed that no maternal Eurasian lineages have been found among Sub-Saharan Africans with a R1- M173 profile.
Haplogroup V88 has the greatest frequency in Africa. It is predominately carried by Chadic speakers, ranges between 2-60% among Central African Niger-Congo speakers (Cruciani et al, 2010). Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010).
.
The phylogeography of R1 in Africa makes it clear that this y-chromosome is spread globally across Africa and includes the genetic structure of diverse African populations including Berber, Chadic, Cushitic, Khoisan,Pygmy, Niger-Congo, Nilo-Saharan and Semitic speaking African populations (Berniell-Lee et al, 2009; Cruciani et al, 2010; Wood et al, 2009). The fact that Dravidians carry the R haplogroup illustrate the recent introduction of R y-chromosome to Eurasia.
Abu-Amero et al (20009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of Ry-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa, especially Cameroon (Coia et al, 2005). Coia et al (2005) has revealed that no maternal Eurasian lineages have been found among Sub-Saharan Africans with a R1- M173 profile.
Haplogroup V88 has the greatest frequency in Africa. It is predominately carried by Chadic speakers, ranges between 2-60% among Central African Niger-Congo speakers (Cruciani et al, 2010). Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010).
 The phylogenetically deep haplogroup R1b is mainly found in West Africa and the Sahel, where the frequency ranges between 5-85% among various Niger Congo speakers (Cruciani et al, 2010). The paternal record of M173 on the African continent illustrates a greater distribution of this y-chromosome among varied African populations than, in Asia.
The greatest diversity of R1b in Africa is highly suggestive of an Africa origin for this male lineage. Archaeological (Lal, 1963), genetic (Winters, 2008;2010a), placenames (Balakrishnan, 2005) and linguistic data group (Aravanan,1979,1980; Upadhyaya, 1976,1979; Winters 1985a,1985b, 1989) linking Africans and Dravidian support the recent demic diffusion of SubSaharan Africans and gene flow from Africa to Eurasia. An early colonization of Eurasia 4kya by Sub-Saharan Africans and Dravidian carriers of R1-M173 is the best scenario to explain the high frequency and widespread geographical distribution of this y-chromosome on the African continent (Winters, 2010c). Given the greatest diversity of R1- M173, this is the most parsimonious model explaining the frequency of R-M173 in Africa.
The phylogenetically deep haplogroup R1b is mainly found in West Africa and the Sahel, where the frequency ranges between 5-85% among various Niger Congo speakers (Cruciani et al, 2010). The paternal record of M173 on the African continent illustrates a greater distribution of this y-chromosome among varied African populations than, in Asia.
The greatest diversity of R1b in Africa is highly suggestive of an Africa origin for this male lineage. Archaeological (Lal, 1963), genetic (Winters, 2008;2010a), placenames (Balakrishnan, 2005) and linguistic data group (Aravanan,1979,1980; Upadhyaya, 1976,1979; Winters 1985a,1985b, 1989) linking Africans and Dravidian support the recent demic diffusion of SubSaharan Africans and gene flow from Africa to Eurasia. An early colonization of Eurasia 4kya by Sub-Saharan Africans and Dravidian carriers of R1-M173 is the best scenario to explain the high frequency and widespread geographical distribution of this y-chromosome on the African continent (Winters, 2010c). Given the greatest diversity of R1- M173, this is the most parsimonious model explaining the frequency of R-M173 in Africa.
 .
In conclusion, the R haplogroup probably originated in Africa. In my paper POSSIBLE AFRICAN ORIGIN OF Y-CHROMOSOME R1-M173 , I argue that the P clade originated in Africa because 1) the age of R-V88 and 2) the widespread nature of R1 in Africa. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
In Africa we find R-M269 and V88. Clearly, R-V88 is older than R-M269 there is no evidence of archaeological evidence of a back migration or haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Sahara into Eurasia from Middle Africa. The diversity of R1 haplogroups in Africa supports the proposition that the R macrohaplogroups originated in Africa, not Eurasia.
.
In conclusion, the R haplogroup probably originated in Africa. In my paper POSSIBLE AFRICAN ORIGIN OF Y-CHROMOSOME R1-M173 , I argue that the P clade originated in Africa because 1) the age of R-V88 and 2) the widespread nature of R1 in Africa. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
In Africa we find R-M269 and V88. Clearly, R-V88 is older than R-M269 there is no evidence of archaeological evidence of a back migration or haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Sahara into Eurasia from Middle Africa. The diversity of R1 haplogroups in Africa supports the proposition that the R macrohaplogroups originated in Africa, not Eurasia.
Haplogroup R-M173 and R-M269 was probably spread among Native Americans, by Pre- Columbian Africans, and African slaves. Controversy surrounds the origin of R1 among Native Americans. Most researchers believe it was introduced by Europeans, because M2many Europeans carry R-M269 .
We have been lied too about Black History. When I was growing up my mother made it clear that we were part Choctaw. So in 1967, I took a survey at my High School: DuSable, in Chicago and found that over 40% of my classmates had Indian heritage. ChoctawAt the time many people laughed at us because they only saw Lakota ad Apache on T.V. Today because of the WWW, there are numerous pictures of Black Native Americans on the Web. Below French artist Alexandre de Batz's renderings of Native American life in colonial Louisiana, such as "Desseins de Sauvages de Plusieurs Nations" ("Drawings of Several Native Americans of Various Nations") from 1735
ChoctawAt the time many people laughed at us because they only saw Lakota ad Apache on T.V. Today because of the WWW, there are numerous pictures of Black Native Americans on the Web. Below French artist Alexandre de Batz's renderings of Native American life in colonial Louisiana, such as "Desseins de Sauvages de Plusieurs Nations" ("Drawings of Several Native Americans of Various Nations") from 1735
 Below a Blackfoot warrior by Karl Bodmer, between 1840 and 1843.The majority of Native Americans in North America carry haplogroup R M173.
Below a Blackfoot warrior by Karl Bodmer, between 1840 and 1843.The majority of Native Americans in North America carry haplogroup R M173. R1-M269 probably originated in Africa. In my paper POSSIBLE AFRICAN ORIGIN OF Y-CHROMOSOME R1-M173 ; I argue that the P clade originated in Africa because 1) the age of R-V88 and 2) the widespread nature of R1 in Africa. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 in Europe is probably 8kya (Balaresque et al, 2010). Y-Chromosome R1b1b2 has high frequencies in England, France, Italy and Germany (Balaresque et al, 2010). Clearly, R-V88 is older than R-M269 .
R1-M269 probably originated in Africa. In my paper POSSIBLE AFRICAN ORIGIN OF Y-CHROMOSOME R1-M173 ; I argue that the P clade originated in Africa because 1) the age of R-V88 and 2) the widespread nature of R1 in Africa. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 in Europe is probably 8kya (Balaresque et al, 2010). Y-Chromosome R1b1b2 has high frequencies in England, France, Italy and Germany (Balaresque et al, 2010). Clearly, R-V88 is older than R-M269 .  This figure from the Gonzalez et al article found that 10 out of 19 subjects in the study carried R1b1-P25 or M269. This is highly significant because it indicates that 53% of the R1 carriers in this study were M269, this finding is further proof of the widespread nature of this so-called Eurasian genes in Africa among populations that have not mated with Europeans.Gonzalez et al proposes a West to East spread for P-25, with a possible entry of this clade into Europe via Gibraltar. There is a variety of R haplogroups in Africa.Finding Africans carrying R-M269, in addition to V88, is nothing new. As early as 2009 R-M269 was found in Africa.
This figure from the Gonzalez et al article found that 10 out of 19 subjects in the study carried R1b1-P25 or M269. This is highly significant because it indicates that 53% of the R1 carriers in this study were M269, this finding is further proof of the widespread nature of this so-called Eurasian genes in Africa among populations that have not mated with Europeans.Gonzalez et al proposes a West to East spread for P-25, with a possible entry of this clade into Europe via Gibraltar. There is a variety of R haplogroups in Africa.Finding Africans carrying R-M269, in addition to V88, is nothing new. As early as 2009 R-M269 was found in Africa.
Berniell-Lee et al (2009) found in their study that 5.2% carried Rb1*. The frequency of among the Bantu ranged from 2-20. The frequency of R1b1 among the Pygmy populations is 5% (Berniell-Lee et al, 2009). The frequency of R1b1 among the Guinea-Bissau populations was 12% (Carvalho et al,2010). Gonzalez et al found a high percent of R-M269 in Guinea.Most Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya)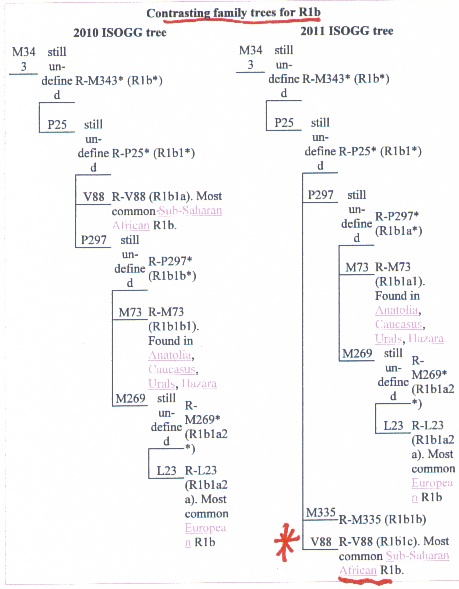 The most recent common ancestor for R1b1b2 in Europe is probably 8kya (Balaresque et al, 2010). Y-Chromosome R1b1b2 has high frequencies in England, France, Italy and Germany (Balaresque et al, 2010) .Clearly, R-V88 dating to 9kya, is older than R-M269 .
In summary the frequency of R-M269 in Africa and age of V88, suggest that R-M269 originated in Africa. The presence of Black Native American communities on the Eastern Seaboard, a region where R1 is found suggest that it was these Black Indians who introduced R1 to America, not Europeans.
The most recent common ancestor for R1b1b2 in Europe is probably 8kya (Balaresque et al, 2010). Y-Chromosome R1b1b2 has high frequencies in England, France, Italy and Germany (Balaresque et al, 2010) .Clearly, R-V88 dating to 9kya, is older than R-M269 .
In summary the frequency of R-M269 in Africa and age of V88, suggest that R-M269 originated in Africa. The presence of Black Native American communities on the Eastern Seaboard, a region where R1 is found suggest that it was these Black Indians who introduced R1 to America, not Europeans.
 Henn et al presents conclusive evidence that African hunter-gatherer (HG) populations share a number of ancestral lineages including B264*; although they are geographically distinct populations situated among agropastorial groups (1). An interesting finding of Henn et al was the discovery of the Eurasian clade R1b1b1a1a among the Khomani San of South Africa (1).Henn et al was surprised by this revelation of R-M269 among this Khoisan population. As a result, he interviewed the carries of R1b1b2a1a, and learned that no members of their families had relations with Europeans. The presence of R lineages among HG populations is not new. Wood et al reported Khoisan carriers of R-M269 (2). Bernielle-Lee et al, in their study of the Baka and Bakola pygmies foud the the R1b1* haplogroup (3). These researchers made it clear that the Baka samples clustered closely to Khoisan samples (3).The most common R haplogroup in Africa is V88. Given the interaction between HG groups and agropastoral groups they live in close proximity too, we would assume that African HG would carry the V88 lineage. Yet, as pointed out above the HG populations carry R-M269 instead of V88 (1-3). The implications of R-M269 among HG populations, and Henn et al’s of shared African HG genome suggest that R-M269 may represent a HG genome. The low frequency of this Eurasian clade among HG populations may not support this conclusion, the distribution of R-M269 among HG populations needs further research into the origins of the R y-chromosome among African populations. References:1. Henn BM, Gignoux CR, Jobin M, Granka JM, Macpherson JM, Kidd JM, Rodríguez-Botigué L, Ramachandran S, Hon L, Brisbin A, Lin AA, Underhill PA, Comas D, Kidd KK, Norman PJ, Parham P, Bustamante CD, Mountain JL, Feldman MW. Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proc Natl Acad Sci U S A. 2011 Mar 29;108(13):5154-62. Epub 2011 Mar 7. http://www.pnas.org/content/108/13/5154.full 2. Wood,E.T., Stover,D.A., Ehret,C., Destro-Bisol,G., Spedini,G., McLeod, H., Louie,L., Bamshad,M., Strassmann,B.I., Soodyall,H., Hammer,M.F. 2005. Contrasting patterns of Y-chromosome and mtDNA variation in Africa:evidence for sex-biased demographic processes. European Journal of Human Genetics, 13:867-876.3. Berniell-Lee G, Calafell F, Bosch E, Heyer E, Sica L, Mouguiama-Daouda P, van der Veen L, Hombert JM, Quintana-Murci L, Comas D. Genetic and demographic implications of the Bantu expansion: insights from human paternal lineages. Mol Biol Evol. 2009 Jul;26(7):1581-9. Epub 2009 Apr 15.http://mbe.oxfordjournals.org/content/26/7/1581.full.pdf .
Henn et al presents conclusive evidence that African hunter-gatherer (HG) populations share a number of ancestral lineages including B264*; although they are geographically distinct populations situated among agropastorial groups (1). An interesting finding of Henn et al was the discovery of the Eurasian clade R1b1b1a1a among the Khomani San of South Africa (1).Henn et al was surprised by this revelation of R-M269 among this Khoisan population. As a result, he interviewed the carries of R1b1b2a1a, and learned that no members of their families had relations with Europeans. The presence of R lineages among HG populations is not new. Wood et al reported Khoisan carriers of R-M269 (2). Bernielle-Lee et al, in their study of the Baka and Bakola pygmies foud the the R1b1* haplogroup (3). These researchers made it clear that the Baka samples clustered closely to Khoisan samples (3).The most common R haplogroup in Africa is V88. Given the interaction between HG groups and agropastoral groups they live in close proximity too, we would assume that African HG would carry the V88 lineage. Yet, as pointed out above the HG populations carry R-M269 instead of V88 (1-3). The implications of R-M269 among HG populations, and Henn et al’s of shared African HG genome suggest that R-M269 may represent a HG genome. The low frequency of this Eurasian clade among HG populations may not support this conclusion, the distribution of R-M269 among HG populations needs further research into the origins of the R y-chromosome among African populations. References:1. Henn BM, Gignoux CR, Jobin M, Granka JM, Macpherson JM, Kidd JM, Rodríguez-Botigué L, Ramachandran S, Hon L, Brisbin A, Lin AA, Underhill PA, Comas D, Kidd KK, Norman PJ, Parham P, Bustamante CD, Mountain JL, Feldman MW. Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proc Natl Acad Sci U S A. 2011 Mar 29;108(13):5154-62. Epub 2011 Mar 7. http://www.pnas.org/content/108/13/5154.full 2. Wood,E.T., Stover,D.A., Ehret,C., Destro-Bisol,G., Spedini,G., McLeod, H., Louie,L., Bamshad,M., Strassmann,B.I., Soodyall,H., Hammer,M.F. 2005. Contrasting patterns of Y-chromosome and mtDNA variation in Africa:evidence for sex-biased demographic processes. European Journal of Human Genetics, 13:867-876.3. Berniell-Lee G, Calafell F, Bosch E, Heyer E, Sica L, Mouguiama-Daouda P, van der Veen L, Hombert JM, Quintana-Murci L, Comas D. Genetic and demographic implications of the Bantu expansion: insights from human paternal lineages. Mol Biol Evol. 2009 Jul;26(7):1581-9. Epub 2009 Apr 15.http://mbe.oxfordjournals.org/content/26/7/1581.full.pdf .
quote:
Genetic and paleoanthropological evidence is in accord that today’s human population is the result of a great demic (demographic and geographic) expansion that began approximately 45,000 to 60,000 y ago in Africa and rapidly resulted in human occupation of almost all of the Earth’s habitable regions. Genomic data from contemporary humans suggest that this expansion was accompanied by a continuous loss of genetic diversity, a result of what is called the “serial founder effect.” In addition to genomic data, the serial founder effect model is now supported by the genetics of human parasites, morphology, and linguistics. This particular population history gave rise to the two defining features of genetic variation in humans: genomes from the substructured populations of Africa retain an exceptional number of unique variants, and there is a dramatic reduction in genetic diversity within populations living outside of Africa. These two patterns are relevant for medical genetic studies mapping genotypes to phenotypes and for inferring the power of natural selection in human history. It should be appreciated that the initial expansion and subsequent serial founder effect were determined by demographic and sociocultural factors associated with hunter-gatherer populations. How do we reconcile this major demic expansion with the population stability that followed for thousands years until the inventions of agriculture? We review advances in understanding the genetic diversity within Africa and the great human expansion out of Africa and offer hypotheses that can help to establish a more synthetic view of modern human evolution.
- human population growth
- hunter-gatherer demography
- molecular evolution
Genetic data indicate that, approximately 45 to 60 kya, a very rapid population expansion occurred outside of Africa, and spread in all directions across the Eurasian continents, eventually populating the entire world. We dub this event the Great Expansion (Fig. 1). The precise location of the exit from Africa, and the relative timing of the southern coastal migration vs. the expansion into northern Eurasia, are still under intense debate (1⇓–3). However, as we discuss here, many parameters of the Great Expansion are now well understood, including the general timing of the exit, the magnitude of the associated bottleneck, and the mode of subsequent expansion. We review the history of the Out of Africa (OOA) expansion, with an emphasis on recent genomic data, and highlight future avenues of research.

Fig. 1.
Ancient dispersal patterns of modern humans during the past 100,000 y. This map highlights demic events that began with a source population in southern Africa 60 to 100 kya and conclude with the settlement of South America approximately 12 to 14 kya. Wide arrows indicate major founder events during the demographic expansion into different continental regions. Colored arcs indicate the putative source for each of these founder events. Thin arrows indicate potential migration paths. Many additional migrations occurred during the Holocene (11).
It is important first to distinguish between the presence of early near-modern humans in the Near East and the very distinct OOA exit associated with the Great Expansion. It is clear that anatomically near-modern humans occupied the Levant (4) during a warm interglacial period 130 to 80 kya, when this region was ecologically similar to northeastern Africa (5). However, current evidence indicates that this near-modern population did not persist in the Near East and was subsequently replaced by Neanderthals during the following glacial period, with little evidence of temporal overlap (5, 6). It is not until at least 50,000 y ago that evidence of behaviorally modern humans occurs in the archaeological record in the Near East. Only after this point do anatomically and behaviorally modern human remains become widespread in Eurasia. It is unclear what precipitated the tremendous population growth associated with this second occupation of the Near East and subsequent dispersal (7); possibly, cultural advances accumulated to a “tipping point” that supported extreme demographic growth (8), or anatomical changes that are not reflected in the paleoanthropological record [e.g., neuroanatomy (9)] occurred in the ancestral population. The geographic range expansions of humans outside of Africa were almost certainly associated with climatic fluctuations (10); however, the Great Expansion was an unprecedented demic success that occurred when the climate remained substantially colder than the previous interglacial. As discussed later, we know that the ancestral population for this expansion was African, but the rate of growth and structure of the ancestral population remain poorly understood.
[...]

Fig. 2.
Schematic of a serial found effect. We illustrate the effect of serial founder events on genetic diversity in the context of the OOA expansion. Colored dots indicate genetic diversity. Each new group outside of Africa represents a sampling of the genetic diversity present in its founder population. The ancestral population in Africa was sufficiently large to build up and retain substantial genetic diversity.
The third assumption is that there have been no dramatic postexpansion bottlenecks that differentially affected populations from which the serial migration began. If the source population for the expansion suffered a severe bottleneck that reduced its genetic diversity, we should see a poorer linear fit to the decline of heterozygosity with distance from Africa, or erroneously assign a population with higher genetic diversity as the source population. It is this third assumption we believe deserves additional consideration.
[...]
Brenna M. Henn,
L. L. Cavalli-Sforzaa,1, and
Marcus W. Feldmanb,2
Edited by C. Owen Lovejoy, Kent State University, Kent, OH, and approved September 25, 2012 (received for review July 19, 2012)
http://www.pnas.org/content/109/44/17758.full
This idea of diversity is correct, but it is probably not due to "genetic drift, isolation and random mutations ". First of all there is no such thing as "random mutation". Genetic drift could have influenced some hypothetical populations, but the diversity among African people is probably due to different origins for each African/Black population (Khoisan, Pygmy, etc) and later mating among the diverse Black populations. Just as there formerly existed homo erectus, homo habilis etc., there was homo australian, homo khoisan, homo pgymy and homo Sub-Saharan African. This view is supported by the skeletal history of anatomically modern humans in Africa.
Thusly, there was more than one out of Africa (OoA) event in which Black people left Africa. The first OoA event was led by homo Australians who left Africa sometime before 100kya, homo Khoisan 45kya, homo pygmy 15kya and finally homo Sub-Saharan Africans 6kya.
This is the only way to explain African diversity. Henn's models for the "Great Expnasion" lack any foundadtion because he is modeling a hypothetical OoA event in which there was a single 'Great Expansion', when there is no evidence that the so-called Caucasian and Mongoloid populations existed anywhere on earth before 6kya. The first Caucasian skeletons do not appear in the archaeological record before 1500BC, Classical Mongoloids (Indonesian,Japanese, Filipino type) around 4000BC, and contemporary East Asian (Chinese, Korean) 1000BC. The late appearance of these populations, and the record of different African populations originating at different times in history provides no support to Henn's 'Great Expansion' theory. It lacks congurence because the theory is based on the idea of the constancy of habitation of the various contemporary populations in the regions they now inhabit, when the historical and archaeological records fail to support any analogy between the present location of populations in the Americas, Eurasia, and even Africa to the populations that inhabited these regions hundreds, let alone thousands of years ago. This lack of analogy is due to population shifts resulting from migrations and war. These factors would have been the major influence on genetic shifts, instead of isolation, genetic drift and genetic mutation.
The fact remains that Naia's DNA was contaminated and Kennewick man carries the African haplogroup X. Moreover Kennewick man is more related to Africans, Andamanese and Melanesians, rather than mongoloid Native Americans craniometrically and genetically.
The Kennewick Man DNA article makes it clear that Kennewick man is recognized as a PaleoAmerican therefore he has negro ancestry. The researchers claim the Kennewick man’s DNA is mainly related to Native Americans living in South America, rather than North America except for the Colville people on the West Coast. The researchers wrote
quote:
“Despite this similarity, Anzick-1 and Kennewick Man have dissimilar genetic affinities to contemporary Native Americans. In particular, we find that Anzick-1 is more closely related to Central/Southern Native Americans than is Kennewick Man (Extended Data Fig. 5). The pattern observed in Kennewick Man is mirrored in the Colville, who also show a high affinity with Southern populations (Fig. 2c), but are most closely related to a neighbouring population in the data set (Stswecem’c; Extended Data Fig. 4c).”
The authors also noted that:
“However, the genetic affinities of Kennewick Man reveal additional complexity in the population history of the Northern lineage. The finding that Kennewick is more closely related to Southern than many Northern Native Americans (Extended Data Fig. 4) suggests the presence of an additional Northern lineage that diverged from the common ancestral population of Anzick-1 and Southern Native Americans (Fig. 3). This branch would include both Colville and other tribes of the Pacific Northwest such as the Stswecem’c, who also appear symmetric to Kennewick with Southern Native Americans (Extended Data Fig. 4).”
The Native Americans on the Pacific coast were a mixture of mongoloid and Pacific Island negro Native Americans.

The Colville tribe which is related to Kennewick man is a Confederation of Indians who did not die of diseases or murdered by whites so they could take their land.
The Colville tribe is the name given to various Christian Native American tribes that lived at Fort Colville. They include Native American groups that were not exterminated by the whites. The twelve Colville bands are the Methow, Okanogan, Arrow Lakes, Sanpoil, Colville, Nespelem, Chelan, Entiat, Moses-Columbia, Wenatchi, Nez Perce, and Palus. These remnants of Pacific coast tribes formerly mixed with the Black Native Americans this is obvious when we look at Ohlone people who lived in missions on the West Coast.

This means that the Colville tribe is admixed with the Black Native American tribes that formerly dominated the Pacific coast.
The authors of the article, attempt to lie about the negro origin of Kennewick man, the multivariate analysis of Kennewick man’s skull does not support their conclusion. The carniometric measurements also confirm the negro origin of Kennewick man. The researchers wrote:
quote:
Although our individual-based craniometric analyses confirm that Kennewick Man tends to be more similar to Polynesian and Ainu peoples than to Native Americans, Kennewick Man’s pattern of craniometric affinity falls well within the range of affinity patterns evaluated for individual Native Americans (Supplementary Information 9). For example, the Arikara from North Dakota (the Native American tribe representing the geographically closest population in Howells’ data set to Kennewick), exhibit with high frequency closest affinities with Polynesians (Supplementary Information 9). Yet, the Arikara have typical Native-American mitochondrial DNA haplogroups30, as does Kennewick Man. We conclude that the currently available number of independent phenetic markers is too small, and within-population craniometric variation too large, to permit reliable reconstruction of the biological population affinities of Kennewick Man.

Arikara
Kennewick man carried mtDNA haplogroup X, this haplogroup is rare among United States Indians. This haplogroup is carried by Africans.
Amerindians carry the X hg. Amerindians and the European hg X are different (Person, 2004). Haplogroup X has also been found throughout Africa (Shimada et al,2007). Shimada et al (2007) believes that X(hX) is of African origin. Amerindian X is different from European hg X, skeletons from Brazil dating between 400-7000 BP have the transition np 16223 ( Martinez-Cruzado, 2001). Transition np 16223 is characteristic of African haplogroups. This suggest that Africans may have taken the X hg to the Americas in ancient times. This transference is supported by the haplogroups carried by Kennewick man.
References:
Brooke Persons, Genetic Analysis and the Peopling of the New World ANT 570, November 9, 2004. http://74.125.95.104/search?q=cache:2g9_ETY1V38J:www.as.ua.edu/ant/bindon/ant570/Papers/Persons.pdf+haplotype+X&hl=en&ct=clnk&cd=4&gl=us
Martinez-Cruzado, J C, Toro-Labrador, G, Ho-Fung, V, Estevez-Montero, M A, Et al (2001). Mitochondrial DNA analysis reveals substanial Native American ancestry in Puerto Rico,Human Biology, Aug 2001
Makoto K. Shimada*, , Karuna Panchapakesan , Sarah A. Tishkoff , Alejandro Q. Nato, Jr* and Jody HeY, Divergent Haplotypes and Human History as Reveaed in a Worldwide Survey of X-Linked DNA Sequence Variation, Molecular Biology and Evolution 2007 24(3):687-698
