

Who are the Kushites? This is a great question because many
nations and tribes called themselves
Kushites, that may have had different names for their own tribe and contemporary nations.
Kushites, that may have had different names for their own tribe and contemporary nations.
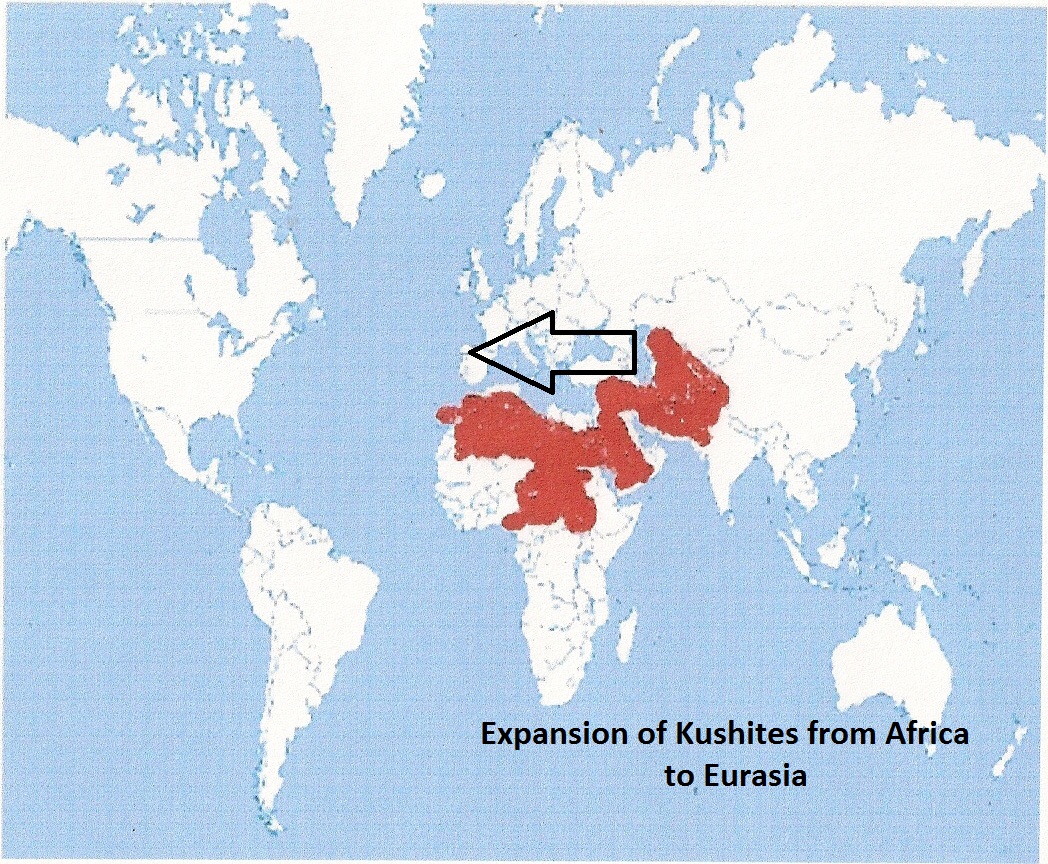
As early as 6000 BC the Kushites had already settled much of the Levant. These Kushites took millet cultivation and cattle from Nabta Playa into Eurasia after 6000 BC.
Narmer was one the first ruler to expand the Kushites into
the Levant and Anatolia. Narmer is attested in Egypt and Canaan. Many serekhs of Narmer have
been found Tel Erani, Arad, 'En Besor, Halif
Terrace/Nahal Tillah .
We know that Narmer was probably a Kushite because of a Clay
bulla (reg. no. G-67.95, Locus 102, B. 1308) from Nahal Tillah. The
inscriptions on the Clay bulla can be read
as ḫЗts.t Kush, ḫ3s.tj "he who
belongs to Kush" or Kushite.

Kushite was just a generic name applied to the people who
belonged to "Kush". But it appears that the Kushites spoke related
but different languages that's why they created lingua franca(s) so the people
would have smooth and consistent means of communication. During the late
Kushite Empire the Kushites in Africa spoke the lingua franca: Meroitic. In
Anatolia the Kushite lingua franca was Nesite.And in Sumer the language of the
Kushites was Sumerian.
As a result, the ancient historians recognized that Kushites
lived in Africa and Asia. Around 800 BC, the Greek poet Homer mentions the
Aethiopians, or Kushites, in the Iliad and the Odyssey.
Homer said that the Kushites were “the most just of men, the favorites of the
Gods”.
To the Greeks and Romans there were two Kush empires, one in
Africa and the other in Asia. Homer alluded to the two Kushite empires when he
wrote in the Odyssey i.23: “a race divided, whom the sloping
rays; the rising and the setting sun surveys”. In the Iliad. i.423,
Homer wrote that Zeus went to Kush to banquet with the
blameless Ethiopians .
In 64 BC, the Greek geographer and historian Strabo stated
in Chapter 1 of Geography that there were two Kush empires -
one in Asia and another in Africa. In addition to Kush in Nubia and Upper
Egypt, some Greco-Roman authors considered their presence in southern Phoenicia
up to Mount Amanus in Syria. Strabo
adds that “if the moderns have confined the appellation Ethiopians to those
only who dwell near Egypt, this must not be allowed to interfere with the
meaning of the ancients.”
The key to knowing who the Kushites were may be explained by
Homer when he wrote in the Odyssey i.23: “a race divided, whom
the sloping rays; the rising and the setting sun surveys”. This suggest that
the Kushites recognized themselves as a “race”.
The Greek word for race is genea ( γενεά, ᾶς, ἡ
): race, family, generation.
Thusly, a Kushite was considered people who belong to same family--men of
the same stock. The idea of the Kushites belonging to the same race implied
that metaphorically, the Kushites in Africa and Asia were like each other in character, endowments, and
pursuits. They were especially identified as great “bowmen” and adventurers.
Ephorus a Greek historian wrote a general history of the
known world. In his histories Ephorus wrote about the Kushites in 405 BC. Ephorus
said that: “The Ethiopians were considered as occupying all the south coasts of
both Asia and Africa,” and adds that “this is an ancient opinion of the of the
Greeks.”

William Chambers, in Information for the People, wrote: “Ephorus
,too (405B.C.)seems to have had a great impression of the power of the
Ethiopians, since he names in the east, the Indians—in the south the
Ethiopians—in the west, the Celts—in the north, the Scythians, as the most
mighty and numerous people of the known earth. Already in Strabo’s time,
however, their ancient power had gone for an indefinite period, and the Negro
states found themselves, after Meroe had ceased to be a religious capital …(p.66)”
Ephorus made a Map of the Ethiopian [Kushite] nations. The Ephorus map, the Classical authors and writers familiar with the Classics like Rawlinson saw the people living in Scythia and Ethiopia (lands below Egypt) as nations founded by Kushites.
As a result, when Strabo wrote:”The north[ern border of Ethiopia/Kush] extends to the remote confines of Scythia and Celtica, and the south to the remote confines of Ethiopia, and the difference between these two extremes is very great ... indeed, they are, in a sense, the antipodes of each other." He was using antipodes as a geographical descriptor, not anthropological identifier of ethnic groups. Strabo’s mention of Ephorus makes it clear he was talking about Kushites who belonged to the same ethnicity, not Black and White ethnic groups.
Strabo also wrote: "[Ephoros] says that if we divide the regions of the heavens and of the earth into four parts, the Indians will occupy that part from which Apeliotes blows, the Ethiopians the part from which Notos blows, the Celts the part on the west, and the Scythians the part from which the north wind blows”. This again alludes to the widespread nature of Kushites around the world in ancient times.
Ephorus’s map of the Kushite nations and dicussion of the Kushites indicates that while there was a geographical antipodes, the people belonged to the “same nationality”.
.
Ephorus made a Map of the Ethiopian [Kushite] nations. The Ephorus map, the Classical authors and writers familiar with the Classics like Rawlinson saw the people living in Scythia and Ethiopia (lands below Egypt) as nations founded by Kushites.
As a result, when Strabo wrote:”The north[ern border of Ethiopia/Kush] extends to the remote confines of Scythia and Celtica, and the south to the remote confines of Ethiopia, and the difference between these two extremes is very great ... indeed, they are, in a sense, the antipodes of each other." He was using antipodes as a geographical descriptor, not anthropological identifier of ethnic groups. Strabo’s mention of Ephorus makes it clear he was talking about Kushites who belonged to the same ethnicity, not Black and White ethnic groups.
Strabo also wrote: "[Ephoros] says that if we divide the regions of the heavens and of the earth into four parts, the Indians will occupy that part from which Apeliotes blows, the Ethiopians the part from which Notos blows, the Celts the part on the west, and the Scythians the part from which the north wind blows”. This again alludes to the widespread nature of Kushites around the world in ancient times.
Ephorus’s map of the Kushite nations and dicussion of the Kushites indicates that while there was a geographical antipodes, the people belonged to the “same nationality”.
.

References:

















{kind=link}